




© Mario Izquierdo
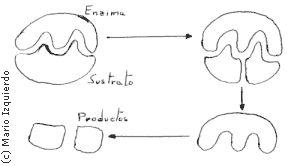
Son en general proteinas y se trata de sustancias catalizadoras que se unen temporalmente a una o más sustancias reaccionantes, produciendo la disminución de la cantidad de energía de activación requrida para la reacción. En la mayor parte de los casos las enzimas se unen a unas sustancias reaccionantes, llamándose entonces sustrato. La unión se realiza por enlaces de hidrógeno, iónicos o entre zonas hidrofóbicas, raramente mediante enlaces covalentes. Las superficies deben quedar cercanas, deben ser a su vez complementarias, para que exista un número importante de este tipo de enlaces débiles. Las enzimas se caracterizan por dos fenómenos:
- La especificidad: Se refiere a que en general una enzima cataliza un tipo de reacción química, o en todo caso varias reacciones en las que los sustratos tengan la misma estructuración básica.
- La inhibición competitiva: Consiste en que si se añade al medio de reacción, una enzima igual al sustrato la enzima se une a ella impidiéndose la reacción. Por ejemplo, la deshidrogenasa succínica, cataliza y produce la oxidación del sulfúrico para dar ácido fumárico, si se añade al medio ácido malónico, la enzima se une a él, inhibiéndose la reacción, por lo tanto, el ácido malónico y el succínico son muy semejantes.
- Factores que afectan a la actuación de las enzimas:
· pH y T°: Cada enzima funciona optimamente a cierta temperatura y pH o entre ciertos límites de estos factores, por ejemplo, pepsina que digiere las proteinas, actúa optimamente entre un pH de 1 a 2, mientras que la tripsina, también enzima proteolítica, es inactiva a un pH comprendido entre 1 y 2, y si es activa a un pH de 8.
Como muchas enzimas son proteinas, su estructura ternaria es fundamental para su funcionamiento, un aumento de temperatura puede destruir los enlaces de hidrógeno, y distorsionar zonas de la estructura ternaria.
Los cambios de pH pueden alterar el estado de ionización de los aminoácidos con carga, como al ácido aspártico o la lisina, son aminoácidos que pueden tener un papel esencial en la unión con el sustrato o la reacción catalítica en si misma.
· Cofactores: Muchas enzimas necesitan la presencia se sustancias, no proteicas adicionales, para su funcionamiento, a estas sustancias se les denomina cofactores, que pueden ser iones como el Mg , que participa en las reacciones en las que se transfiere un grupo fosfato de una molécula a otra, y otros iones como K , Ca , Zn , Cu , Mn , Fe y Na. En otros casos la unión entre aminoácidos e iones mantiene la estructura ternaria en ciertos pliegues y en otros la estructura cuaternaria.
· Coenzimas: Las coenzimas son moléculas orgánicas pequeñas, muchas son vitaminas como es el caso de la riboflavina (B2), tiamina (B1) y nicotinamida. Estas coenzimas pueden reciclarse y volver a ser usadas, por eso el organismo sólo necesita pequeñas cantidades de estas sustancias. Algunas coenzimas pueden unirse covalentemente a la parte proteica de la enzima, dando lugar a un grupo prostético.
- Regulación enzimática:
· Inhibición por retroalimentación o por retracción: Si el producto de una serie de recciones se anula en el interior de una célula, puede inhibir la acción de la primera enzima de la cadena, suspendiéndose la producción. Por otra parte la acumulación de una sustancia dentro de la célula, puede activar específicamente una enzima que desata una serie de reacciones, la sustancia acumulada seria el sustrato inicial, y la acción se conoce con el nombre de activación precursora. La activación precursora, reduce la concentración del sustrato inicial a niveles normales.
Inhibición por retroalimentación
+-------<-------------<--+
| enzima |
a b c d |
Sustrato inicial A ---> B ---> C ---> D ---> E producto final
¦ ¦ ¦ ¦ ^
¦ +-------------+ ¦
¦ metabolitos ¦
¦ intermediarios. ¦
+------>------------->------+
Actuación precursora
Actuación precursora
- Interacción alostérica: En el caso anterior la enzima es regulada por una sustancia diferente del sustrato inicial, el lugar de unión molécula reguladora-enzima, es distinto del lugar de unión sustrato-enzima. La interacción entre puntos reguladores y catalíticos, que se manifiestan por el cambio de forma de la enzima, se conoce como interacción alostérica.
En la inhibición por retroalimentación, la molécula reguladora tiene un efecto alostérico, haciendo disminuir la afinidad de la enzima por el sustrato, mientras que en la activación precursora , aumenta la afinidad de la enzima final de la serie, por su sustrato. La molécula reguladora se llama efector alostérico.
- Inhibición competitiva: Tiene lugar cuando algunos compuestos inhiben la actividad enzimática, ocupando temporalmente el lugar activo de la enzima, entonces el sustrato inicial y el compuesto regulador compiten por el lugar activo, el resultado depende de cuantas moléculas de cada tipo se forman, y en que eslabón de la cadena actúa esta inhibición, por este mecanismo se utilizan varios fármacos que actúan en el tratamiento de infecciones bacterianas en animales, por ejemplo la sulfamida, sustituye al ácido paraminobenzoico (PABA), uniéndose a la enzima que participa en la conversión del ácido PABA, en ácido fólico, éste es imprescindible para las células bacterianas.
- Inhibición no competitiva: Ocurre cuando el inhibidor no tiene porque parecerse al sustrato, y se une a la a la enzima, en lugar distinto al sitio activo. Por ejemplo, el plomo, puede unirse a los grupos sulfuro de aminoácidos, como la cisteina, que forma parte de algunas enzimas, entonces se altera la estructura ternaria de la enzima, inactivándose, la inhibición competitiva y la no competitiva, son reversibles.
- Inhibición irreversible: Las inhibiciones irreversibles son aquellas en las que los inhibidores se fijan al lugar activo de la enzima, o de la desnaturalizan sin que esta puede constituir su estructura ternaria. Por ejemplo, los llamdos gases nerviosos actúan sobre las enzimas que participan en la transmisión del impulso nervioso, de forma que este proceso no puede tener lugar, produciéndose parálisis o muerte.
- Regulación de la fuente: Muchas enzimas hidrolizan los enlaces peptídicos de otras enzimas, por eso en muchos casos la célula sólo produce estas enzimas, al menos en forma activa, cuando lo necesita. Algunas enzimas, entonces se utilizan de forma activa, y en el momento de ser utilizadas se activan, en general, con el concurso de otra enzima, como por ejemplo la pepsina que se sintetiza en las células que revvisten la cavidad interna del estómago, en forma de pepsinógeno, que es inativo y posee una cadena adicional de 42 aminoácidos. El pepsinógeno se transforma en pepsina, perdiendo así esta cadena adicional por la acción de otra enzima. De esta forma se evita que exista pepsina activa en el interior de las células, que podriá hidrolizar proteinas necesarias para esta células.
Otras proteinas potencialmente destructoras, quedan incluidas en lisosomas aislándose del resto de la célula.
Otro tipo de enzimas que existen en el interior de la célula no pueden desplazarse libremente, sino que se encuentran ordenadas especialmente, como las que existen en el interior de los cloroplastos y las mitocondrias.
Se conocen unas 2000 enzimas, algunas de las cuales son muy eficaces, como por ejemplo la anhidrasa carbónica, que actúa en la reacción:
CO2 + H2O <-----------> CO3H2
ácido carbónico.
Cada molécula de enzima permite la producción de 10 moléculas de CO3H2 por segundo. La reacción catalizada por esta enzima es 10 veces más rápida que la no catalizada. En los animales esta reacción es fundamental para la transferencia de CO desde las células hacia el torrente sanguíneo, y de ahí a los pulmones.
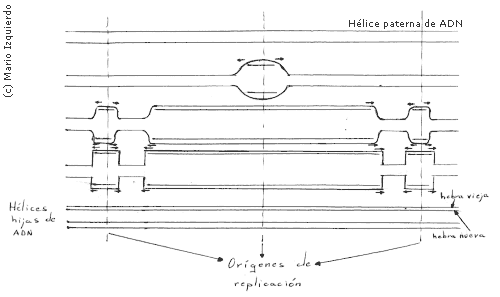
Los organismos vivos deben duplicar con exactitud su ADN antes de cada división celular. La replicación del ADN implica unas velocidades de polimerización de aproximadamente 500 nucleótidos por segundo en las bacterias, y unos 50 nucleótidos por segundo en los mamíferos. El ADN actúa como patrón en el proceso en el que la secuencia de nucleotidos del ADN se copia mediante el apareamiento de las bases complementarias (A=T, G=C). Esto implica el reconocimiento de bases por parte de un nucleotido complementario no polimerizado, y es necesario que las dos hebras de la hélice de ADN se separe para que los grupos dadores y aceptores de enlaces de hidrógeno de cada base, queden libres para que se produzca el apareamiento entre bases. Cada hebra de ADN antiguo se use como patrón para la formación de una nueva hebra, por esto se dice que la replicación del ADN es semiconservativa, es decir, cada molécula de las dos resultantes es una división, hereda una doble hélice con una hebra antigua y una nueva. Existe un región concreta de replicación que se desplaza a lo largo de la hélice paterna de ADN y se conoce con el nombre de horquilla de replicación del ADN, ello es debido a que su forma se parece a Y.
Grupo fosfato
¦ O O
¦ / \ ______¦
H C / \ / \
¦< > BASE ¦¦¦¦¦ BASE < >¦CH
\¦______/ \ / ¦
HEBRA EN O \ / O HEBRA
FORMACION ¦ O ¦ PATRON.
O - P = O O = P - O
5' ¦ ¦
¦ O O
¦ ¦ ¦
3' ¦ O O
¦ / \ ______¦
H C / \ / \
¦< > BASE ¦¦¦¦¦ BASE < >¦CH
\¦______/ \ / ¦
OH \ / O
¦ O ¦
¦ O = P - O
¦ ¦
_____________¦ O O
¦ O / \ ______¦
¦ ¦¦ / \ / \
(1) O - P - O - CH< > BASE ¦¦¦¦¦ BASE < >¦
¦ \¦______/ \ / CH
O OH \ /
O
O O
¦¦ ¦¦
O - P - O - P - (1)
¦ ¦
O O
Esta reacción se cataliza con una enzima que se llama ARN polimerasa. Para esta reacción los sustratos son desoxirribonucleosidos trifosfatos. La reacción se encuentra desplazada por la hidrólisis del pirofosfato que se libera en la reacción de cada nucleotido.
O O O O
¦¦ ¦¦ ¦¦ ¦¦
O - P - O - P - OH + H2O ---> O - P - O - P - OH
¦ ¦ ¦ ¦
O O O O
Los nucleótidos se seleccionan uno a uno mediante el apareamiento de bases, y luego se unen de forma covalente. Para este proceso se necesita una cadena preexistente de polinucleótidos, que se llama cebador de la ADN polimerasa. A medida que la horquilla de replicación se desplaza a lo largo de una molécula de ADN, debido a la orientación antiparalela de las dos hebras, una hebra hija crecería en la dirección 5'-3', la otra en la dirección 3'-5'. Sin embargo utilizando marcaje selectivo, por ejemplo timidina radioactiva, se comprobó que la horquilla de aplicación poseía una estructura asimétrica, una hebra de ADN se sintetiza de forma discontinua en fragmentos de 100 a 200 nucleótidos, que se forman en dirección 5'-3' y que se unen entre sí mediante la acción de otra enzima, que se llama ADN ligasa, y que se activa posteriormente, esta hebra de fragmentos discontinuos se llama hebra retardada y se sintetia después que la otra hebra llamada hebra conductora.
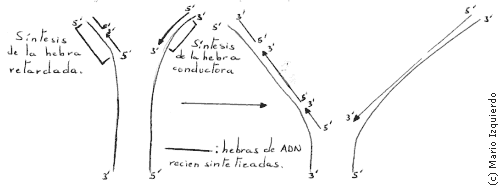
A diferencia de las de ARN polimerasa, que veremos en la síntesis de proteinas, las ADN polimerasas no empiezan una nueva cadena de nucleotido, uniendo entre sí dos nucleosidos trifosfatos, necesitan un extremo 3' hidroxilo (-OH) de cadena de polinucleótidos. Las ADN polimerasas utilizan la actividad de otra enzima que se llama hexonucleasa 3'-5', y que se incorpora a la ADN polimerasa para recortar, mediante hidrólisis, todos los residuos no apareables del extremo de esta hebra cebadora.
En el caso de la hebra retardada se necesita un mecanismo especial para producir los cebadores de bases apareadas que son imprescindibles para iniciar la síntesis de cada uno de los fragmentos de ADN. Entonces una polimerasa que se llama ARN-primasa, utiliza ribonucleosidos trifosfatos para sintetizar cortos fragmentos de ARN, llamado ARN-cebador, de unos 10 nucleótidos de longitud. Estos ARN-cebadores se producen a intervalos sobre la hebra retardada y sobre ellos actúa la ADN polimerasa, formando pequeños fragmentos de ADN que se llaman fragmentos OKAZAKI. La ADN polimerasa, también en este tipo de hebra retardada, continua actuando hasta que se encuentra el ARN-cebador unido al extremo 5' del ADN anterior. Para que los fragmentos de ADN producidos se unan, actúa un sistema especial de reparación del ADN que elimina el ARN-cebador y lo sustituye por ADN.
ARN Síntesis de ARN cebador
cebador 3'por ARN primasa.
3'---------- ------------ <---------------------- 5'
5'----------------------------------------------- 3'
Patrón de la hebra retardada
¦-->¦ADN polimerasa se añade al nue-
¦ ¦vo ARN cebador, inicio síntesis
v ¦fragmentos OKAZAKI.
5' 3'
3'-------- ---- <-------- --- 5'--->3'---------- --- ----------- -- 5'
5'--------------------------- 3' ¦ 5'----------------------------- 3'
+> ADN polimerasa termina fragmento
ADN.
El ARN cebador se elimina¦ ¦
y sustituye por ADN. ¦<--¦
¦
¦
v
3'-------------- -------- --- 5'--->3'-------------------------- -- 5'
5'--------------------------- 3' ¦ 5'----------------------------- 3'
+> ADN ligasa une el nuevo fragmento
de ADN a la cadena en crecimiento.
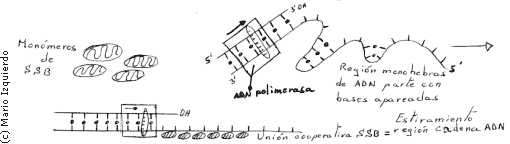
Las ADN polimerasas sólo pueden copiar una molécula de ADN cuando la hebra patrón se halla separado de la doble hélice. Para abrir la hélice son necesarias unas proteinas especiales que son de dos tipos:
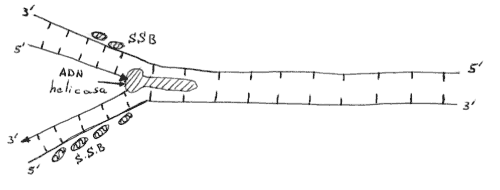
1) Formadas por las proteinas desestabilizadoras de la hélice o proteinas de unión al ADN de una sola hebra o SSB. Estas proteinas se alinean a lo largo de una hebra de ADN extendiendo su estructura y poniendo al descubierto las bases del ADN para su apareamiento. Su presencia perturba las hélices inestables de regiones del ADN con hebras separadas. En la hebra retardada, segmentos de ADN son estirados debido a su unión con proteinas.
2) Proteinas de replicación activadas por ATP. La hidrólisis del ATP puede alterar la forma de una proteina determinando que esta proteina pueda realizar un trabajo mecánico como pasa en el músculo. Proteinas de este tipo, sobre las que actúa el ATP, llamadas ADN helicasas, participan en el desenrrollamiento de las hélices de ADN.
Acción combinada SSB-ADN helicasa
Para que la horquilla de replicación se desplace a una velocidad de 500 nucleótidos por segundo, se necesita que la hélice paterna de ADN gire a 50 revoluciones por segundo:
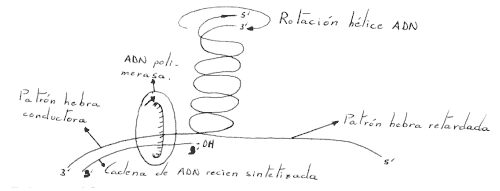
Este problema de arrollamiento, se resuelve por la participación de unos elementos, las ADN-topoisomerasas, que son unas nucleasas reversibles, en primer lugar rompen la cadena de ADN y después la unen de forma covalente.
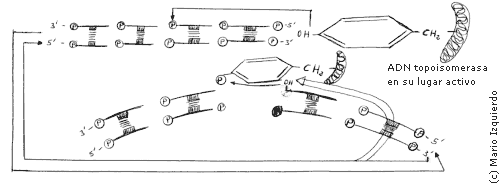
La ADN-topoisomerasa se enlaza covalentemente con el ADN-fosfato, los dos extremos de la doble hélice pueden girar uno respecto al otro, disminuyendo la tensión que hay en la cadena. Se almacena energía en el enlace fosfotiroxina, que permite que la reacción sea reversible y entonces se forma de nuevo el enlace fosfadiester de la cadena de ADN, regenerándose la hélice de ADN y la ADN-topoisomerasa. Estas topoisomerasas que producen la rotura irreversible de una hebra de ADN, se llaman topoisomerasas I, pero además existen las topoisomerasas II, que pueden producir un corte transitorio de la doble hélice de ADN. Esta topoisomerasa II sirve para separar circulos enlazados o nudos de ADN.
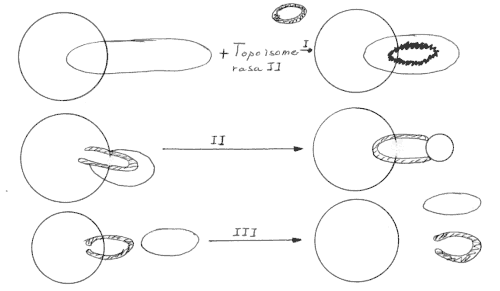
En el primer paso la enzima se une covalentemente a ambas cadenas interrrumpiendo una doble hélice y formando una puerta proteica que corresponde a la enzima, a continuación, la puerta de la topoisomerasa se abre y se cierra, dejando pasar a la segunda hélice, después los dos ADN dúplex circulares se separan y ocurre una reacción inversa a la unión covalente de la enzima restaurándose una doble hélice intacta.
Las horquillas de replicación se originan en una estructura denominada burbuja de replicación, que es una región delimitada en la que las dos hebras de ADN paterno se separan y actúan como patrones para la síntesis de dos nuevas hebras hijas complementarias. Las proteinas de iniciación de la horquilla separarían los pares de bases de la hélice, y sobre una hebra se podría sintetizar ARN-cebador por medio de una ARN-primasa. En las bacterias y algunos virus, las burbujas de replicación se forman en secuencias especiales de ADN, llamados orígenes de replicación, con una longitud de 300 nucleótidos y poseen segmentos especiales que se unen a proteinas especiales para el proceso de iniciación de la horquilla.
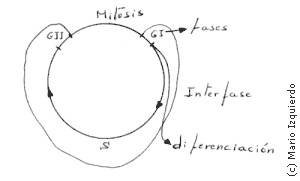
En la fase S, y en relación con la replicación del ADN, existen cinco fenómenos importantes:
• Los orígenes de las horquillas de replicación se activan en grupos, llamados unidades de replicación, cada una de las cuales tiene de 20 a 80 orígenes. Dentro de estas unidades se separa el origen del siguiente por un intervalo de 30.000 a 300.000 pares de bases, por ejemplo cada cromosoma humano posee 150.000.000 de nucleótidos unidos.
• A lo largo de la fase S, se activan nuevos grupos de unidades de replicación hasta que todo el ADN se ha replicado.
• La mayoria de las horquillas de replicación se presentan en parejas, las dos horquillas de un par se mueven en direcciones opuestas, alejándose de un punto común de origen y formándo la burbuja de replicación.
• Las horquillas de una unidad de replicación, terminan su actividad cuando encuentran una horquilla adyacente que se desplaza en dirección opuesta.
• Las horquillas de replicación se mueven a velocidades parecidas en la fase S.
• La fase 'S' dura de 8 a 10 horas.
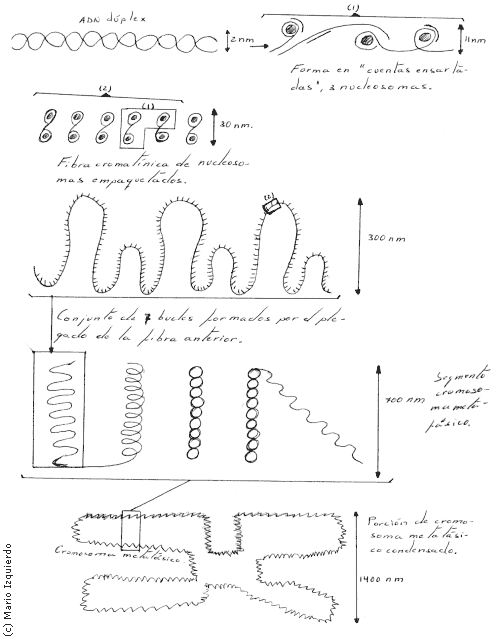
En las células eucarióticas 2 copias de cada una de las cuatro histonas nucleosómicas: H2A, H2B, H3 y H4, dos de estas proteinas se combinan y forman un núcleo proteico con forma de disco compuesta de ocho partes alrededor del cual se arrolla el ADN dúplex del cromosoma, esto lo hace a intervalos de 200 pares de bases, originando así un nucleosoma. Cada bucle tiene una longitud de cromatina que corresponde a decenas de miles de pares de bases. Los bucles actúan como unidades de transcripción del ARN, se condensan y descondensan a medida que se activan y desactivan los genes en la interfase, estas zonas están condensadas y su condensación aumenta en la mitosis, para formar el cromosoma metafásico.
A medida que se va replicando el ADN, se van ensamblando a la cromatina nuevas moléculas de histona que se sintetizan pricipalmente en la fase S, esta síntesis se regula a nivel de transcripción y degradación del ARN-m. Existe un mecanismo de retroalimentación que sigue el nivel de histona libre, y garantiza que la cantidad de histona que se va produciendo es adecuada a la cantidad de ADN que se va sintetizando, esto se demuestra porque si se detiene la síntesis de ADN, el ARN-m que determina la síntesis de las histonas se vuelve inestable y se degrada, sin embargo las histonas son muy estables. Los orígenes de replicación son secuencias específicas del ADN, por tanto las burujas de replicación empiezan en determinados puntos del ADN. La separación entre los orígenes de replicación y los bucles son iguales, por eso es probable que sólo haya un origen de replicación en cada bucle. La localización exacta de un origen de replicación respecto a un gen o unidad de transcripción, determinará como se transmitirán las histonas paternas a los genes hijos. No todos los nucleosomas son iguales y en diferentes regiones génicas existen distintas estructuras cromatínicas, es decir, la localización del origen de replicación respecto a un gen, tiene importantes consecuencias biológicas respecto a la estructura cromatínica de dicho gen. La replicación puede estar controlada de dos formas:
• Por un sistema global, que reconozca específicamente cada banda cromosómica que corresponde a una unidad de replicación, que estaría compuesta por alrededor de 50 bucles y podría descondensar esta banda, haciendo que sus orígenes de replicación resulten disponibles simultáneamente por las proteinas de replicación, que forman las burbujas de replicación.
• Por otro lado, una vez que estas protinas de replicación han encontrado unos cuantos orígenes de replicación de un grupo de replicación resultante, podría perturbar la unidad estrultural cromatínica, estimulando la replicación en los demás orígenes.
Unos factores unidos a la cromatina aseguran que cada región de ADN se replique sólo una vez en cada fase S, la transición de la fase GI a la fase S, está regulada por un iniciador difusible de la síntesis de ADN, la cromatina en GII, en cambio, es incapaz de replicarse porque un factor no difusible, que está fuertemente unido a la cromatina, probablemente modifica la estructura del ADN recien replicado impidiendo que sea de nuevo replicado en ese período.
Entre dos mitosis sucesivas una célula debe duplicar todos sus componentes y por tanto su masa. La mayoría de las proteinas y moléculas distintas, que hay en una célula, se sintetizan de forma contínua a lo largo de toda la interfase, y la capacidad sintetizadora de la célula aumenta a medida que la célula aumenta de tamaño. El ciclo celular avanza siguiendo una serie obligatoria de reacciones secuenciales, un conjunto de controles que actúan durante las primeras fases del ciclo, determinan la masa celular en el momento de la mitosis y debido a esto las células más pequeñas permanecen en GI, hasta que alcanzan el tamaño adecuado. Para controlar el tamaño de la célula a nivel molecular existe una proteina de disparo, que debe acumularse hasta alcanzar un nivel umbral, para que el proceso avance del punto de restricción o GI. Esta proteina podría unirse a puntos cromosómicos, y el proceso avance hacia la fase S, esto exigiría una cantidad de esta proteina superior a la necesaria para unirse a los cromosomas, estos actuarían como inhibidores y sensores de la cantidad de esta proteina.
En la fase GII, algunas de las proteinas que son sintetizadas en esta fase, son esenciales para la mitosis, si se inhibe su síntesis hacia el final de GII, se impide la entrada en la mitosis. Una quinasa soluble (proteinquinasa), que cataliza la fosforilación proteica, podría facilitar el paso de GII a la mitosis, esta enzima fosforilaría las proteinas de la láminar nuclear, lo cual originaría la desintegración de la envoltura nuclear en la mitosis. También esta misma enzima puede fosforilar las moléculas de una histona, que está presente en los nucleosomas, y responsable de su empaquetamiento, es la histona H1, esta fosforilación antes de la mitosis sería la causante de la condensación cromosómica. También a finales de la fase GII, se sintetizan los componentes del huso mitótico.
Los cromosomas replicados en la fase S, se alinean, se separan y desplazan hacia extremos opuestos de la célula (esto es la base de la mitosis), el citoplasma se tiene que segmentar de forma que se asegure que cada célula hija reciba además de un conjunto completo de cromosomas, los elementos y orgánulos citoplasmáticos necesarios (idea básica de la mitosis). Para que tenga lugar la división celular es necesario que haya una maquinaria mitótica, que desde la evolución las células sintetizen grandes cantidades de ADN, incluidos en un número relativamente pequeño de cromosomas. Esta maquinaria asegura una repartición exacta de los cromosomas que se han replicado entre las dos células obtenidas en la división celular. En las células de levadura se produce un error en la separación cromosómica cada 100.000 divisiones celulares.
En esta fase la cromatina que está en forma difusa en la interfase se condensa lentamente, formando cromosomas bien definidos, cuyo número es característico de la especie. Cada cromosoma que se ha duplicado en la fase S o de síntesis, se compone de dos cromatidas, estas dos cromatidas se unen por una estructura llamada centrómero o constricción primaria o pinetocoro. Durante esta fase el nucleolo se descondensa y desaparece progresivamente, los microtúbulos citoplasmáticos que forman parte del citoesqueleto, se disgregan al comienzo de la profase, y constituye una reserva de tubulina. Estas moléculas de tubulina se utilizan en la formación del huso mitótico, que es una estructura fibrilar bipolar que se ensambla fuera del núcleo. El huso mitótico se forma con los centriolos como polos, el par de centriolos que hay originalmente en la célula se replican en un proceso que es inmediatamente anterior a la fase S, dando lugar a otro par de centriolos, en este proceso primero se separan los dos miembros de un par, y a continuación se forma un centriolo fijo en posición perpendicular al antiguo, en su extremo basal. Los centriolos inmaduros presentan nueve microtúbulos sencillos, que sirven de plantilla para el ensamblaje de cada triplete, cada par de centriolos formados se conocen con el nombre de diplosoma, que forma parte de un centro mitótico que constituye el foco de ordenación radial de un conjunto de microtúbulos llamado áster. Los dos ásteres se sitúan inicialmente uno junto a otro, cerca de la membrana nuclear, hacia el final de la profase los microtúbulos polares, que forman hazes, interaccionan entre los dos ásteres y parecen empujar a los dos centros mitóticos, separándolos en la parte externa del núcleo, dando lugar al huso mitótico dipolar.
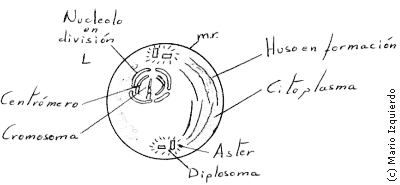
La prometafase se inicia con la desintegración de la envoltura nuclear, que da lugar a fragmentos de membrana semejantes a las vesículas del retículo endoplasmático que son visibles cerca del huso mitótico. En los dos casos de los centrómeros se desarrollan los cinetocoros, que se unen a un conjunto de microtúbulos denominados fibras o microtúbulos cinetocóricos. Estas fibras irradian en direcciones opuestas desde cada lado de cada cromosoma, y entran en interacción con los microtúbulos del huso polar, por esta interacción los cromosomas se desplazan.
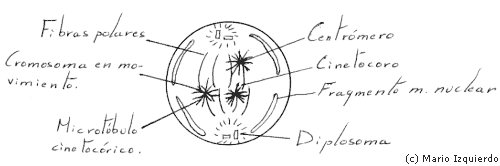
Un huso medio contine 100.000.000 de moléculas de tubulina, que forman microtúbulos, y además existe una reserva de tubulina soluble que está en equilibrio dinámico con los microtúbulos del huso. Los centros que organizan microtúbulos en la célula en división, son los centros mitóticos, que en las células animales se asocian a los centriolos. Se pensaba que estos centriolos eran el centro del ensamblaje del huso, pero en muchas especies incluso en plantas supeiores, se forman husos funcionales en ausencia de centriolos y ásteres, denominados husos anastrales. Además cuando los centriolos se destruyen por un microrrayo LASER, el huso sigue siendo funcional, el centro organizaría los microtúbulos, pero no es esencial para su distribución y ensamblaje en un huso mitótico. Al microscopio electrónico se ha visto que el centro organizador de microtúbulos es una nube de material teñido que está cerca de los polos de los husos astrales y anastrales. Este material aislado in vitro, hace crecer los microtúbulos polares del aster. Es posible que la posición polar de los diplosomas asegure la segregación en las dos células hijas. El cinetocoro al microscopio electrónico es una estructura multiseriada a la que se asocian microtúbulos, que puede ser cinetocórico en los hongos, y de 20 a 40 en el hombre.
En esta fase los cromosomas se disponen de forma que todos los centrómeros estén en un plano, los microtúbulos cinetocóricos son responsables de la orientación de los cromosomas en el ecuador, y perpendicularmente al eje del huso, produce interacción entre las fibras cinetocóricas y el resto del huso, produce la orientación y recolocación de los cromosomas en el plano ecuatorial, dando lugar a la placa metafásica. Este movimiento en los mamíferos dura de 10 a 20 minutos y pertenece a la prometafase. Los diferentes cromosomas giran y oscilan entre los polos, y finalmente un conjunto u otro de fibras polares captura a un cinetocoro, entonces el otro cinetocoro del cromosoma se asocia poco después al otro conjunto de fibras polares. El comportamiento de los cinetocoros se ha estudiado por el uso de la micromanipulación, así se ha comprobado que los cinetocoros no son específicos de un polo del huso, y se puede conseguir que dos conjuntos de fibras cinetocóricas de un mismo cromosoma, vayan al mismo polo. Los cromosomas están estacionarios en la placa metafásica porque las fuerzas dirigidas hacia los polos son iguales y opuestas.
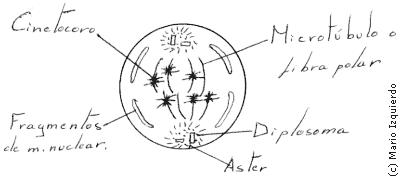
La anafase comienza cuando se separan los cinetocoros que están apareados en cada cromosoma, permitiendo que cada cromátida sea arrastrada lentamente hacia el polo del huso a una velocidad igual a 1 μm/min. Los microtúbulos cinetocóricos se apartan a medida que los cromosomas se acercan a los polos. Las fibras del huso se alargan y los polos se separan más entre si.
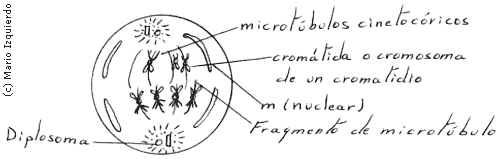
Una posibilidad de explicar el mecanismo que mantiene unidas dos cromátidas hasta la anafase, es que la secuencia que especifica la existencia de un centrómero, puede codificar una señal que bloquea la replicación del ADN durante la fase 'S'. El ADN no replicado del centrómero mantiene unidas las cromátidas, y su replicación posterior constituye el impulso para la separación de la cromátidas. Las fusiones centroméricas se especifican con una secuencia de ADN, con menos de mil pares en la levadura. Estas secuencias se encargan también de la organización de los cinetocoros.
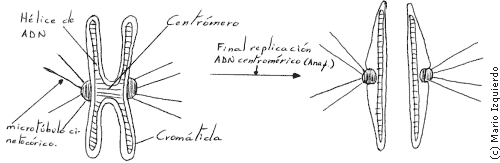
Los microtúbulos polares se superponen en la región del ecuador del huso, estos dos conjuntos se deslizan unos respecto al otro en la zona de superposición durante la anafase, y se alargan mediante la adición de subunidades de tubulina en sus extremos libres, estos microtúbulos polares poseen un extremo de bajo crecimiento, situado debajo de los polos, y alto en el área de superposición. Los microtúbulos cinetocóricos tienen su extremo de rápido crecimiento unido al cinetocoro, tanto los microtúbulos polares como los cinetocóricos tienen la misma polaridad estructural. Una proteina parecida a la dineina, que se encuentra en los cilios y flagelos, y que genera una fuerza de deslizamiento por medio de la hidrólisis de ATP, formando puentes cruzados entre los microtúbulos, y provoca el movimiento entre los dos conjuntos de microtúbulos. En la anafase el movimiento de cada cromátida requiere una energía de hidrólisis de 20 moléculas de ATP.
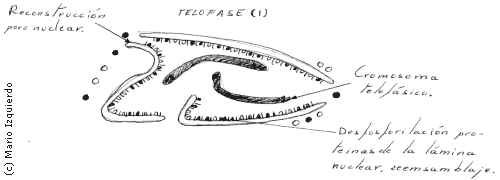
Cuando las cromátidas llegan a los polos, los microtúbulos cinetocóricos desaparecen, las fibras o microtúbulos polares se alargan y se forma una nueva envoltura nuclear alrededor de cada grupo de cromátidas libres. El conjunto de la cromatina que forman los cromosomas se expande y aparecen nucleolos.
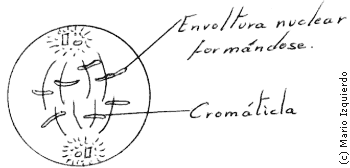
En la disgregación y ensamblaje del complejo de la envoltura nuclear, hay que tener en cuenta dos estructuras: las membranas nucleares externas e internas, que se continúan con las membranas del retículo endoplasmático y la lámina nuclear. La lámina nuclear es una fina estructura compuesta por tres proteinas principales que interactúan con la membrana nuclear interna, cromatina y poros nucleares, en la lámina nuclear de los vertebrados existen tres polipéptidos de peso molecular entre 60.000 y 70.000.
Citoplasma
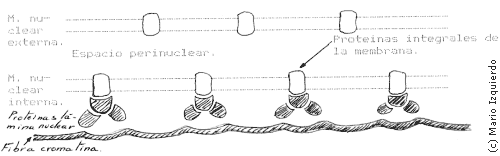
Otra estructura importante que paticipa en la envoltura nuclear son los poros nucleares. Algunas estructuras asociadas a los poros nucleares pueden permanecer asociadas a los cromosomas durante la mitosis. Los fragmentos de la membrana nuclear rodea a cada cromosoma, antes de fusionarse, formando la envoltura nuclear completa.

La cinetocinesis se suele iniciar en la anafase, y continúa a través de la telofase hasta la interfase. En las células animales el primer signo de segmentación es la aparición de un surco, formado por la invaginación de la membrana plasmática, este surco se forma siempre en el mismo plano, que corresponde a la placa metafásica. Si el surco se desplaza en una etapa temprana de la cinetocinesis, desaparece posteriormente y se forma un nuevo surco en el plano ecuatorial del huso, cuando el surco está establecido, la segmentación continúa, aunque el huso mitótico sea eliminado.
Los microfilamentos de actina del citoesqueleto se disgregan en sus subunidades, y los filamentos intermedios, que forman una red que rodea al núcleo interfásico, se alargan durante la mitosis, rodeando a los dos núcleos hijos, y separándose finalmente por la acción del surco de segmentación. La segmentación se hace sobre todo, por la contracción de un anillo, formado principalmente por filamentos de actina, este anillo contráctil se forma al principio de la anafase, junto a la cara interna de la membrana plasmática y conserva el mismo grosor a lo largo de toda la segmentación, entonces se reduce su volumen debido a la pérdida de filamentos, y finalmente se elimina cuando la membrana plasmática se estrecha junto al puente medio, que sirve de conexión entre las dos células hijas, este cuerpo medio contiene restos de microtúbulos polares que quedan empaquetados por material denso de la matriz citoplasmática, y puede persistir durante algunas horas. Puede ocurrir que el anillo contráctil se desarrolle en una posición tal que la segmentación de lugar a dos células asimétricas, esto se puede observar en las primeras fases del desarrollo de algunos embriones, donde existe una división diferencial de las células en zonas distintas.
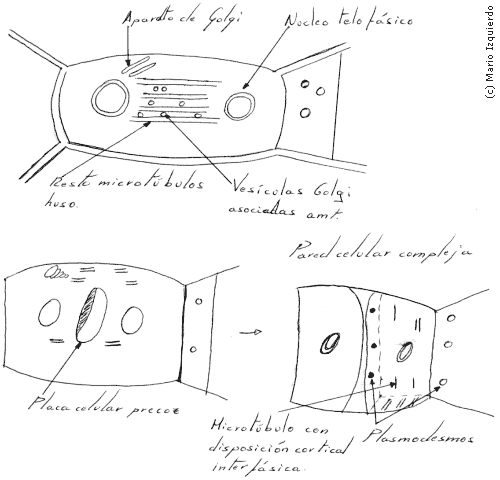
En las células vegetales, como la mayoria, poseen pared celular externa y rígida, en la cinetocinesis el citoplasma se divide desde el interior de la célula, por la construcción de una nueva pared en las células hijas y no por estrangulamiento en un anillo contráctil. El plano de división determina la forma de la planta, la nueva pared, que también se llama placa celular, se forma en relación con los microtúbulos del huso, que constituyen una estructura abierta y cilíndrica, en series paralelas denominadas fragmoplastos.
Unas vesículas de pequeño tamaño, que derivan en gran parte del aparato de Golgi, y llenas de precursores de pared celular, entran en contacto con el fragmoplasto y se transportan por los microtúbulos hasta la región ecuatorial, en esta zona, las vesículas se fusionan y forman una estructura discoidal rodeada de membrana, denominada placa celular precoz. Las moléculas de polisacáridos precursoras, liberadas por las vesículas, se unen entre si en la placa celular precoz y forma pectina, hemicelulosa y otros componentes de la pared celular primaria. El disco se extiende lateralmente hasta llegar a la pared celular original, los microtúbulos del fragmoplasto se disgregan y ensamblan de nuevo en la periferia de la placa celular precoz, y desde allí atraen a un segundo conjunto de vesículas que se fusionan en el ecuador, extendiéndose así la placa. Este proceso se repite hasta que la placa celular llega a la membrana plasmática y separa por completo las dos células. En el interior de la placa se depositan microfibrillas de celulosa, que completan la pared celular. Asociados a las vesículas de la placa en formación, hay elementos del retículo endoplasmático que quedan atrapados a través de la placa, estos elementos se transforman más tarde en plasmodesmos, que son poros complejos que atraviesan la pared celular y permiten la conexión entre el citoplasma de unas células y otras de las plantas.
© Mario Izquierdo, 2024 | Aviso legal | Política de Privacidad